Dijk D Archer S N 2009 Light Sleep and Circadian Rhythms Together Again Plos Biology
Introduction
The low-cal-dark cycle is considered the most important synchronizer of the human circadian pacemaker (Duffy and Wright, 2005) and together with sleep homeostasis, determines slumber propensity (Dijk and Czeisler, 1995). Sources of light bachelor to humans are the natural light-night cycle and "electric" light. Even though the natural light-nighttime cycle is driven by geophysical cycles, exposure to natural light is largely determined by behavioral patterns, e.1000., indoor piece of work (Wright et al., 2013). Exposure to electric light is also linked to behavioral cycles, the about prominent being the sleep-wake cycle, which is frequently dictated past social constraints such as work and school schedules (Wittmann et al., 2006). The interaction of light exposure, behavioral cycles, social constraints and sleep homeostasis has been summarized in mathematical models (Phillips et al., 2010; Skeldon et al., 2017).
Associations betwixt habitual diurnal light exposure, sleep and cyclic outcomes, in the "ecological" dwelling house environment, point that people with early on timing of sleep and daytime activities (early types) have increased light exposure in the early on morning hours, whereas belatedly types take increased lite exposure in the evening hours with a lower aamplitude of the light-dark exposure cycle (Goulet et al., 2007; Emens et al., 2009; Martin et al., 2012; Van der Maren et al., 2018). The duration of bright lite exposure (>thousand lux) is slightly college in early compared to late types (Goulet et al., 2007; Emens et al., 2009), with a greater deviation betwixt the extreme morning and extreme evening types (Emens et al., 2009). A study of individuals with delayed sleep stage confirmed that relative to clock time, their light exposure patterns were shifted to later in the mean solar day compared to controls (intermediate sleep phase) but, relative to cyclic fourth dimension, i.due east., dim calorie-free melatonin onset (DLMO), their low-cal exposure occurred before than in controls (Wilson et al., 2018).
Young individuals unremarkably exhibit a delayed sleep stage (Hagenauer et al., 2009; Gradisar and Crowley, 2013) and consistently demonstrate large differences between slumber timing on weekdays and weekends (Roenneberg et al., 2004; Shochat et al., 2017). This is thought to reverberate the biological shift toward after sleep timing that starts in early puberty and ends around age 20 (Roenneberg et al., 2004; Hagenauer et al., 2009). Developmental changes in the circadian clock, changes in sleep homeostasis, early schoolhouse and work regimes, social forces including peer pressure, reduced daytime light and increased evening low-cal exposure take all been implicated as causal factors (Crowley et al., 2018).
While many field studies examining associations betwixt light, habitual slumber and circadian outcomes and private differences therein, focus on the function of factors such as chronotype, they rarely consider the impact of weekday-weekend, seasonal changes, and changes betwixt standard fourth dimension (ST) and daylight saving time (DST) (Goulet et al., 2007; Emens et al., 2009; Burgess and Molina, 2014; Chinoy et al., 2018).
Seasonal changes dictate differences in day length, which partly determines overall lite exposure including its spectral limerick (Thorne et al., 2009; Stothard et al., 2017) and thereby may affect sleep schedules. Subsequently bed and wake times in wintertime relative to summertime were observed in university students in Norway, where there are large seasonal differences in the daily photoperiod, but not in students from equatorial Republic of ghana where there are no seasonal differences in photoperiod (Friborg et al., 2012). On the other hand, seasonal effects on slumber duration are very small, i.e., in the guild of minutes (Allebrandt et al., 2014) or not statistically pregnant (Lo et al., 2014). In the latter newspaper, a comparison of weekday/weekend sleep schedules across seasons between participants from Singapore (very pocket-size changes in photoperiod, no DST) and Surrey (large changes in photoperiod and DST), indicated that both sleep timing and duration are primarily adamant past social zeitgebers, e.1000., clock time and associated low-cal exposure rather than the natural light-dark cycle.
The impact of the timing of the natural light dark cycle relative to clock fourth dimension has been derived from the report of spontaneous sleep timing (i.e., sleep timing during weekends) in individuals living at different longitudes inside a time zone. People living on the west side of time zones where solar noon (SN) occurs at a afterward clock time, study sleeping later on complimentary days compared to those living on the e side of the time zone (Roenneberg et al., 2007). These observations are in accord with the notion that the human being circadian system is entrained by "sun time" (Roenneberg et al., 2007).
In describing and interpreting changes in slumber timing in relation to calorie-free exposure and its changes across seasons, information technology is important to ascertain the phase reference points used to quantify timing differences. When dawn or sunset are used every bit phase reference points, the timing of social constraints which for most people are stage-locked to clock time, change across seasons. Employing time elapsed from the middle of the dark period has been suggested to exist a more informative reference point (Daan et al., 2002), or similarly, time elapsed from the heart of the light menstruation, i.e., SN. SN, unlike dawn and dusk, does not depend strongly on photoperiod. SN remains at much the same clock time throughout the year in countries without DST. However, in countries with DST, the alter from ST to DST is associated with an abrupt shift of 1 h for SN (and dawn and dusk) relative to clock time and associated social constraints. As an culling to SN, dawn or dusk, the weighted midpoint of bodily light exposure can be used as a phase reference signal, or the unabridged time series of calorie-free tin be entered into mathematical models aimed to assess the effects of lite, using the model predicted minimum of the cyclic oscillator every bit the reference point (Woelders et al., 2017). A prediction that follows from the notion that the human circadian timing organization is entrained to sun time is that spontaneous sleep timing, every bit a proxy for circadian phase and quantified in local clock time, will be one h later during DST compared to ST. Alternatively, it may be argued that the man circadian timing organisation is entrained to social schedules and or actual low-cal exposure, which is a combination of natural and electric lite, in which case spontaneous sleep timing will be associated with clock fourth dimension or the timing of light exposure.
Ecological studies to engagement take all the same to focus on the inter-individual variability of the timing of slumber and activity, and its association with light exposure on weekdays versus weekends and beyond seasons. Thus the aim of the present study in undergraduate academy students in the United Kingdom was to assess sleep timing (i.east., bed, wake, and mid-sleep times), and timing of the midpoint of daily activity, on weekdays and weekends and in belatedly autumn (ST) and in tardily spring (DST), relative to clock time, to SN, and to the timing of the weighted midpoint of light exposure, i.e., the fourth dimension at which 50% of the total daily light exposure has accumulated (fifty%). Specifically we assessed (i) 24 h light exposure (LE), level and timing of 50% LE, by season (autumn/spring), twenty-four hours-type (weekday/weekend) and bedtime category (early on/intermediate/belatedly); (ii) Bed, wake, mid-sleep and weighted midpoint (fifty%) of daily activity times by season, mean solar day-blazon and bedtime category, relative to clock time, SN, and fifty% LE; and (3) nosotros compared our experimental data to the predictions of the furnishings of light according to a mathematical model for the circadian and homeostatic regulation of slumber (Skeldon et al., 2017).
Materials and Methods
Ideals and Participants
The research protocol was approved by the University of Surrey Ethics Committee and was conducted in accordance with the principles of the Declaration of Helsinki. Written informed consent was obtained from all the participants prior to starting any study related procedures. Twenty-three healthy young men and women were recruited into the protocol after responding to local advertisements for participation in the written report, and undergoing an initial phone screening. Inclusion criteria called for full-time university students residing in university dormitories or off- campus housing. Exclusion criteria (all ascertained with validated health and sleep questionnaires) included working night shifts, having or being currently treated for a sleep disorder or for depression, having ophthalmologic or other neurological abnormalities, astute or chronic disease, taking medications on a chronic ground, particularly medications affecting the key nervous system, alcohol intake >14 units per week on average and daily consumption of more than than 4 cups of caffeinated beverages (e.g., java, tea, cola) over the preceding i month. Of the 20-3 participants, ii did not complete the measurements and two were removed due to incomplete information, leaving 19 participants (mean age ± 1SD: 18.88 ± 0.83 years) who completed the study and were included in the analyses presented here.
Report Design and Procedures
In this repeated measures observational study design, all measurements were collected during the late fall of 2022 (Nov to Dec – starting at least 22 days after the change to ST, hereafter, autumn) and once again in the late bound (April–May, 2022 – starting at to the lowest degree 29 days later on the change to DST, hereafter, spring). Each assessment segment consisted of 3-weeks (during the semester) of continuous monitoring of daily habitual lite exposure and rest-activeness patterns, self-assessed bed and wake times and evening sleepiness levels (these data are non discussed here). The 24-h light exposure and residue-activity cycles were recorded via actigraphy. Self-reported daily bedtimes and wake times were recorded with sleep diaries. Furthermore, seasonal differences in natural illuminance were recorded, with an actiwatch encased in a clear container, for two weeks during each assessment segment outdoors at 51°13′36″Northward 0°38′36″West, a dark-green belt area without electrical calorie-free sources in its vicinity.
Assessments of Daily Light Exposure, Rest-Activeness Patterns, and Sleep Patterns
Low-cal exposure and activity
This was measured with Actigraphy technology (Actiwatch-50, CamNtech Ltd.). The Actiwatch-L is a wrist-worn ambulatory device designed for studies in naturalistic settings that features continuous objective recording of white lite illuminance in units of lux, and activity data (wrist movements at minute resolution) that is scored as epochs of sleep and wake, concomitantly. We computed daily light exposure and action levels described below from these data.
Sleep
The Karolinska Sleep Diary was used for assessing daily sleep patterns. This self-report slumber diary provides information on cocky-selected daily bed and wake-times.
Deriving Light, Activeness and Sleep Measures
Light
Daily (24-h) light exposure (24-h LE)
These are the daily (midnight to midnight) hourly averages computed individually for every participant. The data were log-transformed for analyses because the circadian effectiveness of light is linearly related to the log of lite intensity over a wide range of intensities (Zeitzer et al., 2000).
Solar noon (SN)
The clock time at which the sun crossed the local meridian and reached the highest betoken. SN information was collected for all participants per written report twenty-four hours from: https://world wide web.timeanddate.com/sunday/united kingdom of great britain and northern ireland/guildford.
Midpoint of light exposure (50% LE)
This is the time at which 50% of the daily calorie-free exposure had accumulated. This was computed equally one-half the cumulative light exposure (using log-transformed light levels) between wake time and bedtime, and was done for each study 24-hour interval for every participant.
Activity
Daily (24-h) activity levels
These are the daily (midnight to midnight) hourly averages computed individually for every participant.
Midpoint of action (50% deed)
This is the fourth dimension at which 50% of the daily cumulative activity was reached. 50% of the daily action level was computed as half the cumulative activity level between wake time and bedtime, and was done for each written report day individually for every participant.
Sleep
Bedtime (BT)
These are self-reported bedtimes recorded in daily slumber diaries. Those from Sunday through Thursday were designated as weekday times, while those from Friday through Saturday were designated as weekend times.
Wake time (WT)
These are self-reported wake times recorded in daily sleep diaries. Those from Monday through Fri were designated as weekday times, while those from Saturday through Dominicus were designated as weekend times.
Sleep duration (SD)
This was computed every bit the time elapsed between BT and WT.
Mid-sleep (MS)
This midpoint of the time betwixt slumber onset (sleep diary) and wake time (WT) (slumber diary), and was computed for each written report day individually for every subject. Mid-sleep (MS) has been proposed to exist a proxy for circadian phase (Kantermann and Burgess, 2017).
BT category (BTcat)
Participants were divided to BT categories based on their weekday bedtimes during the autumn using the post-obit cutoffs: Early (E):<=23:30 (northward = 8); Intermediate (I): between 23:30 and 01:00 (n = 4), and Late: (L):>=01:00 (n = seven).
The sleep measures were aligned to local clock time, to SN and to l% LE for the analyses.
Statistical Analyses
All the data were analyzed using the statistical package SAS ix.4 (SAS Institute Inc., Cary, NC, Usa). For daily light and activeness levels and for BT, WT, MS and fifty% human activity (aligned to local clock fourth dimension, SN and 50% LE) we used general linear mixed model (Procedure MIXED, SAS 9.iv) ANOVA, with two levels each for season (spring and fall) and 24-hour interval-type (weekday and weekend). The ML estimation method was used with a compound symmetry covariance structure for the subject event. For all of the above variables, we then added 3 levels for the betwixt subjects factor BTcat (described to a higher place) to the mixed model. The ML estimation method was used with compound symmetry covariance structure for the bailiwick consequence and unstructured covariance for the within-subjects effect. To ascertain individual variability we computed Intra Class Correlations (ICC) using the covariance parameter estimates from the mixed model ANOVA. ICC = CS (compound symmetry estimate) subject/(CS subject field + rest). The significance of associations between the various measures were ascertained with Pearson's correlations computed using Process Corr (SAS ix.4; SAS Establish Inc., Cary, NC, United States) and testing the goose egg hypothesis of significance.
Model Simulations
Calorie-free profiles during fall and bound and weekends and weekdays, averaged across participants, were used as input to the mathematical model reported in Skeldon et al. (2017). The model includes the entrainment of the circadian pacemaker by calorie-free and the homeostatic and circadian regulation of sleep. This model as well describes the neuronal flip-flop switch that results in consolidated sleep and wake states. Switching between states is driven by circadian rhythmicity and slumber homeostasis. The mathematical model is an adaptation of the model of Phillips et al. (2010), incorporating social constraints and calibrated to brand quantitative predictions of sleep-wake timing given realistic light profiles.
Physiological parameters for model simulations
The model includes three physiological parameters that are peculiarly relevant for determining individual differences in slumber duration and timing. These are: intrinsic circadian menstruation, the rate of homeostatic rising during wake and circadian amplitude. As default values, we have the intrinsic circadian catamenia as 24.2 h, consequent with the measured mean value (Duffy et al., 2011). We have selected the rate of homeostatic ascension during wake and circadian aamplitude to give a spontaneous sleep elapsing of 8.9 h and mid-sleep time of 05:00 for an intrinsic cyclic period of 24.ii h and the boilerplate low-cal profile used in Skeldon et al. (2017). These friction match the average slumber duration and timing for age xix years reported in Roenneberg et al. (2004) and Roenneberg (2013). The rationale for fitting age based on homeostatic rise during wake and circadian aamplitude is discussed in Skeldon et al. (2016).
Social constraints
Sleep timing is determined past the interplay of social and physiological factors. Given the differences between measured wake and bed times betwixt weekdays and weekends, we make the assumption that wake during the calendar week is determined by social factors and require that the model wake fourth dimension occurs no later than the measured wake fourth dimension. Nosotros and then predict wake times at the weekend and sleep time, cyclic stage and sleepiness for all days.
Results
Overall Effects of Day-Blazon and Season
The natural low-cal-dark cycle and average low-cal exposure, activity and sleep timing during weekdays and weekends during fall and jump are displayed in Effigy 1.
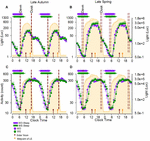
Figure one. 24-h Activeness and light exposure profiles during weekdays and weekend in the belatedly fall and late spring. The average 24-h light (A,B) and activity (C,D) profiles of participants during the ii seasons are shown equally double plots in the figure. The expanse plot in the background of all four panels represents an average 24-h profile of natural illuminance during the two seasons. The horizontal bars testify the average weekday (purple) and weekend (green) sleep, respectively, with the left and right error bars indicating the standard mistake of bed and wake times, respectively. The dashed reference lines indicate the dawn and dusk ranges during the study period. The brown and orangish arrows stand for the average SN and the boilerplate midpoint of light exposure times. The horizontal-axis represents clock time and the left vertical -axis represents light counts (A,B) and activity levels (C,D). In both panels the right vertical axis represents light levels corresponding to the natural illuminance.
Low-cal Exposure (24-h and fifty% LE) and Activity Levels (24-h)
Participants were exposed to lite for a menstruum much longer than the natural photoperiod on both weekdays and weekends, in both autumn and jump (Figures 1A,B). While the daily (24-h) low-cal exposure level did not differ betwixt 24-hour interval-types, information technology was higher in spring than in autumn (Table 1, and Figures 1A,B). SN was plain later in spring (DST) (12:58, range 12:58–13:01) than in fall (ST) (eleven:50, range 11:46–11:55). The midpoint of calorie-free exposure (50% LE) ranged from early afternoon to evening in both seasons (Figure 2A). Relative to clock fourth dimension, 50% LE occurred afterward SN (Figure 1) and significantly afterward on weekends compared to weekdays (p = 0.01), with no seasonal differences (Table 1). Relative to SN, 50% LE occurred earlier in spring than in fall on both weekdays and weekends (Tabular array 1). Like light exposure, participants' activity menses was longer than the natural photoperiod on weekdays and weekends, particularly in fall (Figures 1C,D). The average daily activity level (24-h) was college on weekdays compared to weekends in both seasons (Table 1).

Table i. Averages, standard errors of the hateful (SEs) and intra-class correlation coefficients (ICCs) of 24-h log lux (and retransformed), 24-h activity counts, and timing of l% of individual daily low-cal exposure (50% LE).

Figure 2. Midpoint of light exposure (fifty% LE) and mid-slumber time relative to SN and clock fourth dimension. (A) shows individual 50% LE times relative to SN in late autumn and belatedly spring. (B) shows individual mid-slumber times during weekends, weekdays in both seasons relative to the SN. (C) shows the human relationship between fifty% LE time and mid-sleep fourth dimension, relative to clock fourth dimension on averaged weekdays and weekends and in both seasons. Mid-sleep time is significantly correlated with the 50% LE exposure time, which explains 42% of the variance in mid-sleep time
Timing of Sleep and Activity Relative to Clock Time, SN and 50% LE
The timing of sleep and activity occurred later on weekends than on weekdays, but with some seasonal differences.
Relative to clock time, there was a main effect of day-blazon (p < 0.01) on sleep and activity times, with BT, WT, MS, and 50% act occurring later on weekends compared to weekdays (Table two and Figure i). There were no seasonal effects on these measures, with the exception of WT which occurred significantly after on weekends in autumn compared to weekends in spring (day-type by season interaction: p = 0.03). There was a main result of day-type (p < 0.01) on SD in that it was longer on weekends in both seasons (Table 2).

Tabular array 2. Averages, standard errors of the mean (SEs) and intra-form correlation coefficients (ICCs) of sleep and activity measures by Clocktime.
Relative to SN (Table 3), at that place were main effects of day-type (p < 0.01) and flavor (p < 0.0001) on BT, WT, MS, and 50% act, in that they occurred significantly after weekends than on weekdays and earlier in bound (DST) than in autumn (ST). For WT at that place was a twenty-four hours-type by season interaction, so that weekday-weekend differences were larger in fall than in spring.

Tabular array iii. Averages and standard errors of the mean (SEs) of sleep timing (BT, WT, MS) and 50% action later SN.
Relative to 50% LE (Tabular array iv), there was a significant event of 24-hour interval-blazon (p < 0.05) just on WT and MS, both of which were significantly later on weekends than on weekdays. MS, in both seasons, was distributed between 03:00–08:00 (Effigy 2B). A significant correlation between MS and 50% LE explained 42% of the variance in MS, in that a later MS was associated with a later fifty% LE (Figure 2C).

Tabular array four. Averages and standard errors of the mean (SEs) of sleep timing (BT, WT, MS) and l% activity after l% Calorie-free EXPOSURE (50% LE).
Individual Differences in Sleep Timing: Stability Beyond Seasons and Day-Type
Individual differences in sleep timing (BT and WT) are illustrated in Effigy iii. In all four panels, participants are ordered according to bedtimes on weekdays in autumn. The height panels show the within-private consistency of BT across day-blazon and flavour, while the bottom panels show this in WT. The order of BT (primeval to latest, across participants) is not maintained in WT, indicating that BT is non a skillful predictor of WT. This was confirmed by a correlation analyses, in which BT-WT correlations by season and mean solar day-type did not accomplish significance (Supplementary Table S3). However, highly significant correlations for mean and median BT, WT, and MS between weekday and weekend by season (Supplementary Tabular array S1), and between fall and spring by day-type (Supplementary Table S2) and the intraclass correlations (Table 2) indicate that the BT and WT patterns within individuals are consistent across twenty-four hours-type and season.
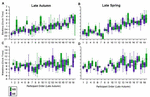
Figure 3. Inter individual differences and intra-individual stability in bed and wake times during weekdays and weekend in the tardily autumn and late spring. The daily bedtimes of participants during both seasons are shown as boxplots in the two upper panels (A,B) while the corresponding wake-times are shown in the ii lower panels (C,D). The purple boxplots stand for weekdays and the green boxplots the weekend days. Participants in all four panels are ordered from earliest to latest weekday autumn bedtimes.
Figure four shows light levels in two individual participants, one with early on BT (A,B) and 1 with late BT (C,D) on weekdays and weekends in autumn (A,C) and in spring (B,D). Photoperiods clearly occur at a later clock time in the belatedly BT participant than in the early BT participant.
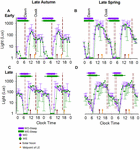
Effigy 4. Weekday/weekend 24-h light exposure profiles and sleep timing during belatedly autumn and belatedly spring in a representative early and a representative tardily bedtime participant. The boilerplate 24-h light profiles of a participant with early (A,B) and belatedly (C,D) sleep timing during both seasons are shown every bit double plots in the effigy. The horizontal confined show the boilerplate weekday (purple) and weekend (dark-green) sleep, respectively, with the left and right mistake bars indicating the standard deviations of bed and wake times, respectively. The dashed reference lines betoken the dawn and dusk ranges during the study period. The brown and orangish arrows correspond the average SN and the average midpoint of light exposure times. The horizontal-centrality represents clock time.
Given this individuality, we analyzed light, sleep and activity timing measures across three bedtime categories (BTcat: early, intermediate and late sleepers) based on their weekday bedtimes in autumn (see cutoffs in section "Materials and Methods"). These measures were all analyzed relative to clock time, SN and 50% LE (Supplementary Tables S4–S6).
Calorie-free, Sleep and Activity in the Three Bedtime Categories (BTcat)
Daily Low-cal Exposure (24-h and 50%) and Activeness (24-h)
There was a meaning main effect of BTcat (F ( 2 , 64 ) = half-dozen.94, p = 0.0019) on the daily 24-h LE exposure (in log lux), in that light levels (mean ± SE) were higher in the early: (1.02 ± 0.04) and belatedly (1.00 ± 0.04) compared to the intermediate (0.77 ± 0.06) types. Effigy 5A summarizes the seasonal and day-type differences in 50% LE, in the three bedtime categories, by clock time and past hours after SN. At that place were meaning master effects of Btcat on the 50% LE, in that it occurred at a later on clock fourth dimension (F ( 2 , 64 ) = four.87, p = 0.0108) and at a afterward time compared to SN (F ( ii , 64 ) = 4.57, p = 0.0140) in the tardily types (hateful ± SE for clock time: sixteen:14 ± 0:14 and for hours afterwards SN: 3.81 ± 0.24) than in the early types (mean ± SE for clock fourth dimension: 15:22 ± 0:xiv and for hours after SN: 2.84 ± 0.23). Note that consistent with Figure 5A, relative to SN, the l% LE occurred earlier in leap than in fall both on weekdays and weekends, but relative to clock time it does not. For 24-h activity, no principal effects and no interactions were constitute. 24-h light exposure and activity profiles in the three bedtime categories are shown in Supplementary Figures S1, S2, respectively.
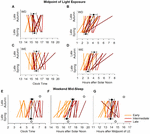
Effigy 5. Midpoint of low-cal exposure (50% LE) and weekend mid-sleep time in the late autumn and late jump. Upper panels show the individual and boilerplate weekday (A,B) and weekend (C,D) fifty% LE times relative to clock fourth dimension (A,C) and SN (B,D). Lower panels prove the private and average weekend mid-sleep times during the late fall and late spring relative to clock time (Due east), solar noon (F) and midpoint of light exposure (G). In all the panels, individuals are color coded for early on, intermediate and late bedtime categories. The filled symbols with mistake bars represent the boilerplate times with between subjects variance (SEM). The open symbols (lower panels) represent the model predicted average mid-sleep times.
Timing of Sleep and Activity Relative to Clock Time, SN and 50% LE
The results of the analyses of slumber timing measures (BT, WT, MS) and 50% act by BTcat relative to clock time, SN and 50% LE are shown in Supplementary Tables S4–S6, respectively. Overall, master furnishings for BTcat were observed for the timing of sleep and activeness (with the exception of WT and 50% act relative to fifty% LE) such that timing measures occurred at a later time from the early to the late BTcat (run across pairwise comparisons for BTcat beyond season and solar day-blazon, and BTcat within season in the Tables). The primary effects of season and twenty-four hour period-type were in accordance with the pattern observed in Tables two–4 (in general, later times on weekends compared to weekdays and earlier in the leap than in autumn).
Figure 5B summarizes seasonal differences in MS during the weekend, for the three bedtime categories, by clock fourth dimension, hours after SN, and hours later 50% LE. Consequent with the in a higher place results, relative to SN the weekend MS occurred before in jump than in autumn, simply relative to clock fourth dimension and l% LE, MS was unchanged across seasons.
Simulating the Effects of Calorie-free Exposure on the Timing of Sleep the Timing of the Maximum of Circadian Slumber Propensity
Nosotros adjacent explored how current models of entrainment of the human cyclic timing system respond to the observed light exposure patterns in the three BTcat. Effigy 6A shows predicted slumber timing, and the timing of the minimum of circadian wake propensity, across the calendar week for the calorie-free exposure patterns measured in spring and autumn for the intermediate bedtime category. The combination of the social constraints on wake time during the week and light exposure results in entrainment to a weekly pattern, with bedtime shifting slightly earlier Monday to Thursday equally a result of increasing slumber debt. On Friday, in spite of a continued rise in slumber debt, bedtime is later than on Thursday. This is the result of entrainment to a weekly schedule leading to a pocket-size aamplitude weekly oscillation in circadian timing with the circadian minimum of the wake propensity rhythm occurring at the earliest on Thursday nighttime and at the latest on Saturday night. And so although sleep debt is greater on Fri than on Thursday, circadian rhythmicity is afterward in "anticipation" of the weekend with the net result of afterward sleep timing on Friday than Thursday. BT and WT are predicted to be later on the weekend and particularly then during fall. The clock time of the circadian minimum of wake propensity is relatively stable across the calendar week occurring ∼0.v h afterward weekends. More noticeable is that the cyclic minimum occurs several hours before WT on weekends but close to WT on weekdays. This effect is more pronounced in autumn than in spring.

Effigy 6. Model predicted weekday and weekend sleep times for early, intermediate and tardily types in the late autumn and late spring. Raster plots (A) for sleep timing in late autumn (dark greyness bars) and tardily spring (light grayness bars) for intermediate types. The left/right paw colored arrows indicate weekday wake time during spring/autumn. (B,C) show hateful computed sleep timing for weekday and weekend for early, intermediate and late groups during the tardily fall (dark gray bars) and tardily spring (light gray bars). For comparison, the measured values are also shown (colored bars). For all panels, the time of the computed circadian minimum wake propensity is indicated by the orange circles.
Panels (B) and (C) show a detailed comparison between average BT and WT measured in the field to average BT and WT and the timing of the circadian wake propensity minimum predicted by the model, for weekdays and weekends. WT for weekdays were determined past social constraints, as well in the simulations. For leap, BT as predicted to within 1 h for all three BTcat during both weekdays and weekends and WT during weekends were predicted to inside 1.5 h. The predicted timing of the minimum of circadian wake propensity was several hours earlier WT in all three BTcat during both weekdays and weekends. For autumn, slumber and wake timing were within 2 h, with the model predicting later BT and later weekend WT than measured. The circadian wake propensity minimum was located very close to WT during weekdays for all three BTcat.
Discussion
Few ecological studies have followed light exposure, activity, and sleep patterns in immature undergraduate students in their natural surround for continuous periods of time, i.due east., iii weeks, repeated over ii seasons of the year (autumn and spring), including weekdays and weekends. Comparing inter-individual variability of slumber patterns beyond seasons and day-type by distinct reference points aims to disentangle the effects of biological, social and environmental factors in sleep regulation. Comparison differences in daily light exposure caused by DST, too as two light exposure parameters (SN and timing of fifty% LE), which are independent of each other and of DST, allowed the states to test the hypothesis that human sleep follows sunday time. This study can exist considered a natural experiment that takes reward of a widely institutionalized social norm (i.e., DST), and examines its association with slumber timing. Such natural experiments are considered to be of high ecological validity. Our findings provide a comprehensive assessment of individual stability and variability in these factors, and demonstrate that individual timing of slumber and action in a modern surround with electrical lighting, largely conforms to clock time and actual light exposure (which is the combination of natural and electrical light exposure) rather than to sun time indexed by the timing of SN.
Light Exposure
Meridian lite levels in the current study (around 600 in the spring and 200 lux in fall) autumn inside the range of actigraphy calorie-free data (50–1000 lux) from ecological studies conducted at similar latitudes (45–51°N) (Goulet et al., 2007; Emens et al., 2009; Thorne et al., 2009; Martin et al., 2012; Van der Maren et al., 2018). Higher light levels have been reported in studies of young adults in the Rocky Mountains in Colorado (twoscore°North), with averaged peak light levels of >2000 lux in the summer (Wright et al., 2013) and winter (Stothard et al., 2017), and extreme seasonal differences have been reported in a study of day workers in Kiruna, Sweden (68°N), with mean peak lite levels during midday close to 2000 lux in summertime, and around 110 lux in winter (Lowden et al., 2018). In a written report of rubber tappers in the Amazon (x°39″ Due south), peak median lite levels were 1200–1600 lux on work and free days, respectively (Moreno et al., 2015).
In the present study, overall 24-h light exposure was higher in bound than in autumn just did not differ between weekdays and weekends. Similar to our findings, in a study assessing weekly light exposure across seasons (wintertime and summer) and day-types (workdays and weekends) in young working adults in Chicago (41°88′Northward), the overall low-cal exposure was lower in winter (autumn ST in our study) compared to summer (spring DST in our study), the photoperiod occurred later in the solar day on weekends than on weekdays, and light levels were higher in summertime than in winter on weekdays and weekends during the early on evening hours, likely due to the longer natural photoperiod (Crowley et al., 2015).
Inter-Individual Variability and Stability in Sleep, Activity and Lite Exposure
Several individual aspects of sleep timing were stable beyond seasons and weekday vs. weekends, in accordance with a previous study which besides the midpoint of sleep also analyzed stability of melatonin phase (Kantermann and Eastman, 2018).
In the electric current study, stability of private differences, as assessed by intra-class correlations (ICCs), were highest for bedtime, about one-half the magnitude for wake time, somewhat lower for mid low-cal exposure and minimal for overall 24-h light exposure and activity (run into Tables ane, ii), indicating that inside individuals, bedtime is the most consistent timing parameter. Indeed, weekday-weekend and autumn-spring bedtime correlations were highly meaning, notwithstanding bedtimes and wake times were not correlated. Furthermore, categorization of individuals as early on, intermediate and late based on autumn weekday bedtime predicted all timing parameters every bit expected; however, furnishings were weaker for wake time and the midpoint of activity and disappeared altogether for these timing parameters when assessed relative to the individual midpoint of light exposure. Findings suggest that bedtime in young individuals is largely tuned by individual preference (or biological determination), whereas wake time is more tuned to social constraints and environmental time cues. Similarly, in a person-centered assay of several sleep timing and sleep continuity indicators of senior high schoolhouse students, bedtime showed the highest outcome size for differences between phenotype classifications (Shochat et al., 2017). In dissimilarity, findings from a study of sleep in 3 preindustrial societies (Yetish et al., 2015) showed greater intra-individual variability in sleep onset than in sleep beginning. These contrary findings may reflect differences in the relative contributions of social constraints and environmental fourth dimension cues to sleep patterns in preindustrial vs. modern 24-hour interval societies (specifically hither, college students). A corollary is that individual sleep duration is better predicted past individual differences in bedtime rather than wake fourth dimension in the preindustrial social club (Yetish et al., 2015), and is meliorate predicted by wake fourth dimension in young university students, when environmental factors and social constraints are held abiding. Still, the availability of electrical evening light has been associated with after bedtime, longer slumber latency and shorter sleep elapsing beyond individual differences in bedtime (Wright et al., 2013; Moreno et al., 2015).
Slumber and Activity Timing Relative to Clock Time, SN and 50% LE
Relative to clock time, sleep and activity timing variables were afterwards, and sleep duration longer, on weekends compared to weekdays. Previous studies take reported later bed and wake times on weekends compared to weekends in adolescents (e.g., Tzischinsky and Shochat, 2011; Shochat et al., 2017) and young adults (due east.g., Lo et al., 2014; Lowden et al., 2018). Our findings of no seasonal differences in sleep timing (during either weekdays or weekends) but significant day-type effects when measured relative to clock fourth dimension are in line with other studies (Lo et al., 2014; Crowley et al., 2015). This, together with our finding of significant seasonal effects on sleep timing when measured relative to SN, confirms that social norms and constraints and associated calorie-free exposure rather than sunday fourth dimension decide observed sleep and activeness regulation. Although in our statistical analyses we used SN as the phase marker for sun time, information technology is articulate that using dawn and sunset would have led to a like conclusion, i.eastward., slumber timing is not phase-locked to either dawn or dusk (run across Figure 1). Thus, the average dusk to bedtime intervals in fall (7-h: 56-min, and 8-h: 34-min for weekdays and weekends, respectively) are about twice those in spring (iii-h: 22-min, and 3-h: 56-min), and increase from early to belatedly bedtime categories. Similarly, dawn to wake time intervals in jump (4-h and 14-min, and 4-h and 50-min) are about twice those in fall (1-h and 59-min, and 2-h and 36-min), and increase from early on to late bedtime categories. Information technology is evident that individual differences and social constraints override the seasonal differences in the timing of SN, sunset and dawn in determining sleep timing.
Seasonal effects on sleep timing may be axiomatic only in higher latitudes (e.1000., Friborg et al., 2014; Lowden et al., 2018). For instance, in a study in Sweden (68°N) which examined sleep timing, activity, light and daytime behavioral measures in wintertime and summertime on work days and days off, seasonal effects were found for all slumber timing variables (sleep onset, beginning and mid-sleep), with later sleep timing and shorter sleep elapsing in wintertime compared to summer (Lowden et al., 2018). In another study of young adults in Kingdom of norway (69 Due north, 39 E), sleep timing and chronotype measures were delayed in December compared to September and March (Friborg et al., 2014).
Withal, flavor interacted with differences in bedtime category and mostly showed gradually afterward sleep timing measures from early to intermediate to late bedtime categories. Thus, the range of bedtimes was larger in leap than in autumn, and both wake time and mid-sleep in autumn were earlier for the early on category only no differences were found between the intermediate and belatedly categories. These findings warrant further investigation of the subtle dynamics of light exposure in autumn versus spring and their effects on individual differences in sleep timing.
When expressed equally "hours later on" SN, sleep and activity timing variables were consistently later in autumn than in spring and, as for clock time, were later on on weekends than on weekdays. Season interacted with mean solar day-blazon for wake time only, showing that later wake fourth dimension in autumn than in spring was more pronounced on weekends (about 1–h and forty-k) than on weekdays (ane-h but), and that weekday-weekend differences were larger in fall (1-h and xx-min) than in spring (45-min). Similar season by bedtime category interactions were observed for the sleep timing variables as described relative to clock time.
The seasonal shift in sleep timing relative to SN (subsequently in the bound due to DST) reflects an alignment with clock time and the meaning affect of social factors rather than sun time in driving sleep timing in young higher students. These findings practice not support the notion that the human cyclic timing system entrains to sunday time equally derived from an assay of seasonal changes in self-reported sleep timing (Allebrandt et al., 2014) and the differences in sleep timing beyond a time zone (Roenneberg et al., 2007). Furthermore, greater weekday-weekend differences in wake time in autumn compared to spring appears to contradict findings (Kantermann et al., 2007) suggesting that sleep timing on free days as assessed by self-report, is subsequently relative to dawn in bound (DST) than in fall (ST) due to difficulty in tracking sunday fourth dimension during DST. Conversely, larger differences in sleep timing from early to belatedly bedtime categories in spring compared to autumn in our study may lend some back up to this claim. The consequences of DST shifts are notwithstanding under contend (Lindenberger et al., 2018; Roenneberg et al., 2019; Skeldon and Dijk, 2019), withal our findings provide piddling support to the suggestion that individuals and particularly late types experience more difficulty adjusting to social constraints (clock time) in spring than in winter, due to the longer photoperiod, or to DST, or to both.
No seasonal differences were found for timing variables relative to the midpoint of light exposure suggesting that actual lite exposure is more of import than sun time. The midpoint of light exposure, but not SN, was significantly correlated with mid-sleep beyond season and 24-hour interval-type, and the average weekday midpoint of light exposure significantly correlated with average weekend mid-sleep (r = 0.46, p = 0.049), suggesting that bodily light exposure is a strong determinant of sleep timing (Woelders et al., 2017). Findings emphasize the importance of collecting private light measurements, rather than measurements from satellites or ground stations, which may atomic number 82 to the wrong conclusion that light does not touch sleep timing (Porcheret et al., 2018). Weekday-weekend differences were constitute for wake fourth dimension and mid-slumber, and season interacted with bedtime category similarly to clock time and SN reference points, with larger early to late timing intervals in spring than in fall. Notably, although our findings show that individually determined daily light exposure, which includes both natural and electric light, is more strongly associated with sleep timing than simply sun time, they do not imply causality. Indeed, these associations may be unidirectional, bidirectional, or may involve yet a third variable, e.1000., social norms and schedules. Yet even though light exposure may be driven by (rather than driving) the slumber-wake cycle, which in turn may exist driven by social constraints, this light exposure will have an impact on the cyclic pacemaker, as nosotros (Skeldon et al., 2017) and others (Wright et al., 2013; Stothard et al., 2017) have shown.
Model Simulation
The model predicts that differences in low-cal exposure patterns, independent of other physiological differences, are sufficient to predict afterwards sleep timing and more social jetlag for late types than intermediate types and for intermediate types than early types. This relative ordering remains, fifty-fifty if the imposed social constraint of wake fourth dimension on weekdays is removed.
The fact that the predictions for the autumn are worse suggests that either societal factors override physiological cues or that the model is overly sensitive to the reduced mean solar day light during the autumn. Less lite during the day will increase the relative importance of light during the evening, and we accept not sought to have account of the high degree of individual variability in the sensitivity of the circadian system to the effects of evening light (Phillips et al., 2019).
Stage was not measured in the field, but the model predicts that the minimum of the circadian wake propensity rhythm occurs closer to wake time on weekdays than on weekends. This effect is more than pronounced in the autumn than during the spring suggesting that there are seasonal differences in circadian alignment. The prediction that the minimum occurs very close to wake fourth dimension during the autumn suggests wake fourth dimension is during the biological night, making it especially hard to get up.
Written report Limitations
As in most studies, light exposure was measured from a wrist worn device and to what extent this accurately reflects retinal light exposure remains unclear. An underestimation of lite exposure is particularly a business organisation in the fall, due to colder temperature and long sleeves. In improver, in this study no information on the spectral composition of 24-h calorie-free exposure, which has been shown to vary across seasons (Thorne et al., 2009), was nerveless. As in other studies (e.grand., Kantermann et al., 2007), we have used slumber timing on weekends as a proxy for cyclic stage. No other accustomed marker of cyclic phase such every bit melatonin or core trunk temperature was recorded. This is particularly relevant because the simulation suggests major changes in the phase relationship between sleep timing and cyclic rhythmicity in detail in the autumn (see Figure 6). The sample size is small, however, individual activeness and light profiles over 24-h (total number of 24-h periods: 715) were collected continuously, at a 1-min charge per unit, for two three-calendar week periods, in a within subject area pattern, i.e., in each participant data were obtained in both seasons, thereby greatly increasing the statistical ability. Few studies have collected such a wealth of information using clothing technology in an ecological setting. Findings may not generalize beyond college students, withal, the same statement may be fabricated for whatsoever other segment of the population. Importantly, it is probable that the daily schedules of young college students are less constrained compared to those of immature adults who exercise non attend college and adults in later years, due to growing work and family responsibilities, and thus their slumber and activity patterns are less confounded. Indeed, population based survey findings that computed mid-sleep on gratis days as a proxy for chronotype, similarly to the present study, take shown that variability in chronotype reaches its peak at ages 20–24 (Fischer et al., 2017), which may reflect not only age-related changes in bio-regulatory processes of circadian stage, but also less constrained slumber schedules on free days as reported by immature adults. Finally, additional seasonal differences such as temperature, which has been shown to affect sleep both in preindustrial (Yetish et al., 2015) and modern day (Hashizaki et al., 2018) societies, was not monitored here; however, in the United kingdom of great britain and northern ireland, differences in temperature are minimal between fall and bound. Thus average 24 h outdoor temperatures for autumn and spring were viii.eight∘C and 10.8∘C, respectively and since nigh individuals spend nearly of their time indoors, the seasonal differences in temperature exposure are likely to accept been even smaller.
Decision
Our findings provide a comprehensive description of the complex interplay between environmental and individual factors that determine sleep timing in immature adults. By relating slumber timing to distinct reference points, nosotros demonstrate that beyond private differences, sleep timing largely conforms to clock fourth dimension and to bodily light exposure rather than to dominicus fourth dimension. The model simulations suggest that current models for the effects of low-cal on the human circadian timing arrangement perform satisfactory during spring just not so during autumn atmospheric condition and imply that assessment of circadian phase markers may provide additional insights into seasonal changes in sleep-wake regulation. Findings may inform the ongoing scientific and socio-political fence regarding the implementation of DST and its consequences on human sleep and health.
Data Availability
The datasets generated for this study are bachelor on request to the respective author.
Ethics Statement
The research protocol received a favorable opinion from the University of Surrey Ethics Committee and was conducted in accordance with the principles of the Declaration of Helsinki. Written informed consent was obtained from all the participants prior to starting whatever study related procedures.
Author Contributions
TS, NS, AS, and D-JD conceptualized the study design and protocol. NS and SF collected and candy the data. NS, PH, TS, AS, and D-JD analyzed and interpreted the data. NS, Every bit, and TS prepared the figures and tables. TS, NS, Equally, and D-JD composed the manuscript.
Funding
This study was funded by the Surrey Sleep Enquiry Centre, with no additional external funding. Independent researcher funds (TS) from the University of Haifa were received for statistical analysis and open access publication fees.
Conflict of Interest Argument
The authors declare that the research was conducted in the absenteeism of whatsoever commercial or financial relationships that could be construed as a potential disharmonize of interest.
Acknowledgments
We thank the report participants.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2019.00882/full#supplementary-textile
Effigy S1 | 24 h light exposure profiles of the early on, intermediate and late types during weekdays and weekend in both seasons. The boilerplate 24 h light exposure profiles of the early (A,B), intermediate (C,D), and tardily (Eastward,F) participants during the late fall and late spring are shown as double plots in the figure. The horizontal bars testify the average weekday (purple) and weekend (green) sleep, respectively, with the left and right mistake bars indicating the standard error of bed and wake times, respectively. The dashed reference lines indicate the dawn and dusk ranges during the autumn and spring. The blue and green arrows represent the average SN and midpoint of light exposure times. The horizontal-axis represents clock hr and the left vertical-centrality lite levels (lower panels).
Figure S2 | 24 h activity profiles of the early, intermediate and belatedly types during weekdays and weekend in both seasons. The average 24 h action profiles of the early (A,B), intermediate (C,D), and late (E,F) participants during the late autumn and late jump are shown as double plots in the effigy. The horizontal bars testify the average weekday (majestic) and weekend (green) sleep, respectively, with the left and right error confined indicating the standard error of bed and wake times, respectively. The dashed reference lines indicate the dawn and sunset ranges during the autumn and spring. The blue and green arrows represent the average SN and midpoint of low-cal exposure times. The horizontal-centrality represents clock hour and the left vertical-axis activity levels (lower panels).
TABLE S1 | Correlations between weekdays and weekends for mean/median BT, WT and MS times past season.
TABLE S2 | Correlations betwixt fall and jump for mean/median BT, WT, and MS times by day-type.
Table S3 | Correlations between mean/median BT and WT on weekdays and weekends and in autumn and leap.
TABLE S4 | Averages, standard errors of the mean (SEs) of BT, WT, MS, and fifty% act by season, day type, and BT category (early on, intermediate, belatedly) relative to clock time.
Tabular array S5 | Averages, standard errors of the mean (SEs) of BT, WT, MS and 50% act, by season, day type, and BT category (early, intermediate, late) relative to solar apex (SN).
Tabular array S6 | Averages, standard errors of the mean (SEs) of BT, WT, MS and fifty% act, by season, twenty-four hours blazon, and BT category (early, intermediate, late) afterward 50% light exposure (LE).
References
Allebrandt, K. 5., Teder-Laving, M., Kantermann, T., Peters, A., Campbell, H., Rudan, I., et al. (2014). Chronotype and sleep elapsing: the influence of season of assessment. Chronobiol. Int. 31, 731–740. doi: ten.3109/07420528.2014.901347
PubMed Abstruse | CrossRef Full Text | Google Scholar
Chinoy, E. D., Duffy, J. F., and Czeisler, C. A. (2018). Unrestricted evening use of calorie-free-emitting tablet computers delays self-selected bedtime and disrupts circadian timing and alertness. Physiol. Rep. six:e13692. doi: 10.14814/phy2.13692
PubMed Abstract | CrossRef Full Text | Google Scholar
Crowley, S. J., Molina, T. A., and Burgess, H. J. (2015). A week in the life of total-time office workers: piece of work day and weekend low-cal exposure in summertime and winter. Appl. Ergon. 46, 193–200. doi: 10.1016/j.apergo.2014.08.006
PubMed Abstruse | CrossRef Full Text | Google Scholar
Crowley, Southward. J., Wolfson, A. R., Tarokh, L., and Carskadon, M. A. (2018). An update on adolescent sleep: new evidence informing the perfect storm model. J. Adolesc. 67, 55–65. doi: ten.1016/j.adolescence.2018.06.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Dijk, D., and Czeisler, C. (1995). Contribution of the cyclic pacemaker and the sleep homeostat to sleep propensity, sleep structure, electroencephalographic slow waves, and sleep spindle activity in humans. J. Neurosci. 15, 3526–3538. doi: 10.1523/JNEUROSCI.15-05-03526.1995
PubMed Abstruse | CrossRef Full Text | Google Scholar
Duffy, J. F., Cain, Due south. West., Chang, A.-M., Phillips, A. J. K., Munch, M. Y., Gronfier, C., et al. (2011). Sex difference in the near-24-60 minutes intrinsic flow of the man circadian timing organization. Proc. Natl. Acad. Sci. The statesA. 108, 15602–15608. doi: 10.1073/pnas.1010666108
PubMed Abstract | CrossRef Full Text | Google Scholar
Emens, J. S., Yuhas, K., Rough, J., Kochar, Northward., Peters, D., and Lewy, A. J. (2009). Phase angle of entrainment in morn and evening-types nether naturalistic conditions. Chronobiol. Int. 26, 474–493. doi: 10.1080/07420520902821077
PubMed Abstract | CrossRef Full Text | Google Scholar
Fischer, D., Lombardi, D. A., Marucci-Wellman, H., and Roenneberg, T. (2017). Chronotypes in the US-Influence of age and sex. PLoS 1 12:e0178782. doi: 10.1371/journal.pone.0178782
PubMed Abstract | CrossRef Full Text | Google Scholar
Friborg, O., Bjorvaten, B., Amponsah, B., and Pallesen, South. (2012). Associations betwixt seasonal variations in day length (photoperiod), sleep timing, sleep quality and mood: a comparison between Ghana (5∘) and Norway (69∘). J. Sleep Res. 21, 176–184. doi: ten.1111/j.1365-2869.2011.00982.x
PubMed Abstruse | CrossRef Full Text | Google Scholar
Friborg, O., Rosenvinge, J. H., Wynn, R., and Gradisar, M. (2014). Slumber timing, chronotype, mood, and beliefs at an arctic latitude (69∘Northward). Sleep Med. xv, 798–807. doi: x.1016/j.slumber.2014.03.014
PubMed Abstract | CrossRef Full Text | Google Scholar
Goulet, G., Mongrain, V., Desrosiers, C., Paquet, J., and Dumont, M. (2007). Daily calorie-free exposure in morning-type and evening-type individuals. J. Biol. Rhythms 22, 151–158. doi: x.1177/0748730406297780
PubMed Abstract | CrossRef Full Text | Google Scholar
Hagenauer, M. H., Perryman, J. I., Lee, T. M., and Carskadon, M. A. (2009). Adolescent changes in the homeostatic and circadian regulation of sleep. Dev. Neurosci. 31, 276–284. doi: 10.1159/000216538
PubMed Abstract | CrossRef Full Text | Google Scholar
Hashizaki, Yard., Nakajima, H., Shiga, T., Tsutsumi, M., and Kume, One thousand. (2018). A longitudinal large-calibration objective slumber data analysis revealed a seasonal sleep variation in the Japanese population. Chronobiol. Int. 35, 933–945. doi: 10.1080/07420528.2018.1443118
PubMed Abstruse | CrossRef Full Text | Google Scholar
Kantermann, T., and Eastman, C. I. (2018). Circadian phase, circadian period and chronotype are reproducible over months. Chronobiol. Int. 35, 280–288. doi: 10.1080/07420528.2017.1400979
PubMed Abstract | CrossRef Full Text | Google Scholar
Kantermann, T., Juda, 1000., Merrow, Grand., and Roenneberg, T. (2007). The human circadian clock's seasonal adjustment is disrupted by daylight saving time. Curr. Biol. 17, 1996–2000. doi: 10.1016/j.cub.2007.10.025
PubMed Abstract | CrossRef Full Text | Google Scholar
Lindenberger, L. K., Ackermann, H., and Parzeller, M. (2018). The controversial fence about daylight saving time (DST)—results of a retrospective forensic autopsy study in Frankfurt/Main (Germany) over x years (2006–2015). Int. J. Legal Med. 133, 1259–1265. doi: 10.1007/s00414-018-1960-z
PubMed Abstract | CrossRef Full Text | Google Scholar
Lo, J. C., Leong, R. L. F., Loh, K. K., Dijk, D. J., and Chee, Yard. W. L. (2014). Young adults' slumber duration on piece of work days: differences betwixt E and West. Front end. Neurol. 5:81. doi: 10.3389/fneur.2014.00081
PubMed Abstract | CrossRef Full Text | Google Scholar
Lowden, A., Lemos, N., Gonçalves, B., Öztürk, G., Louzada, F., Pedrazzoli, Thousand., et al. (2018). Delayed sleep in wintertime related to natural daylight exposure among chill 24-hour interval workers. Clocks Sleep i, 105–116. doi: 10.3390/clockssleep1010010
CrossRef Full Text | Google Scholar
Martin, J. Southward., Hébert, Yard., Ledoux, E., Gaudreault, M., and Laberge, L. (2012). Relationship of chronotype to sleep, Lite exposure, and work-related fatigue in student workers. Chronobiol. Int. 29, 295–304. doi: 10.3109/07420528.2011.653656
PubMed Abstract | CrossRef Full Text | Google Scholar
Moreno, C. R. C., Vasconcelos, S., Marqueze, Eastward. C., Lowden, A., Middleton, B., Fischer, F. M., et al. (2015). Sleep patterns in Amazon condom tappers with and without electric low-cal at home. Sci. Rep. 5:14074. doi: 10.1038/srep14074
PubMed Abstruse | CrossRef Total Text | Google Scholar
Phillips, A. J. K., Vidafar, P., Burns, A. C., McGlashan, E. Thou., Anderson, C., Rajaratnam, S. M. W., et al. (2019). Loftier sensitivity and interindividual variability in the response of the human circadian arrangement to evening low-cal. Proc. Natl. Acad. Sci. U.Due south.A. 116, 12019–12024. doi: 10.1073/pnas.1901824116
PubMed Abstruse | CrossRef Total Text | Google Scholar
Phillips, A. J. K. K., Chen, P. Y., and Robinson, P. A. (2010). Probing the mechanisms of chronotype using quantitative modeling. J. Biol. Rhythms 25, 217–227. doi: x.1177/0748730410369208
PubMed Abstract | CrossRef Total Text | Google Scholar
Porcheret, K., Wald, L., Fritschi, 50., Gerkema, M., Gordijn, M., Merrrow, Grand., et al. (2018). Chronotype and environmental light exposure in a student population. Chronobiol. Int. 35, 1365–1374. doi: 10.1080/07420528.2018.1482556
PubMed Abstruse | CrossRef Total Text | Google Scholar
Roenneberg, T., Kuehnle, T., Pramstaller, P. P., Ricken, J., Havel, Thou., Guth, A., et al. (2004). A marker for the finish of boyhood. Curr. Biol. 14, R1038–R1039. doi: 10.1016/j.cub.2004.11.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Roenneberg, T., Wirz-Justice, A., Skene, D. J., Ancoli-Israel, S., Wright, K. P., Dijk, D.-J., et al. (2019). Why should we cancel daylight saving time? J. Biol. Rhythms 34, 227–230. doi: x.1177/0748730419854197
PubMed Abstruse | CrossRef Full Text | Google Scholar
Shochat, T., Barker, D. H., Sharkey, K. Thou., Van Reen, E., Roane, B. M., and Carskadon, Thou. A. (2017). An arroyo to understanding sleep and depressed mood in adolescents: person-centred sleep nomenclature. J. Slumber Res. 26, 709–717. doi: 10.1111/jsr.12550
PubMed Abstract | CrossRef Total Text | Google Scholar
Skeldon, A. C., Derks, G., and Dijk, D.-J. J. (2016). Modelling changes in sleep timing and duration across the lifespan: changes in circadian rhythmicity or sleep homeostasis? Sleep Med. Rev. 28, 96–107. doi: 10.1016/j.smrv.2015.05.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Skeldon, A. C., Phillips, A. J. Thousand., and Dijk, D.-J. (2017). The effects of self-selected calorie-free-dark cycles and social constraints on human slumber and circadian timing: a modeling approach. Sci. Rep. 7:45158. doi: 10.1038/srep45158
PubMed Abstruse | CrossRef Full Text | Google Scholar
Stothard, E. R., McHill, A. W., Depner, C. Thou., Birks, B. R., Moehlman, T. M., Ritchie, H. K., et al. (2017). Circadian entrainment to the natural calorie-free-nighttime cycle across seasons and the weekend. Curr. Biol. 27, 508–513. doi: ten.1016/j.cub.2016.12.041
PubMed Abstruse | CrossRef Total Text | Google Scholar
Thorne, H. C., Jones, K. H., Peters, S. P., Archer, S. North., and Dijk, D. J. (2009). Daily and seasonal variation in the spectral composition of lite exposure in humans. Chronobiol. Int. 26, 854–866. doi: x.1080/07420520903044315
PubMed Abstract | CrossRef Full Text | Google Scholar
Tzischinsky, O., and Shochat, T. (2011). Eveningness, slumber patterns, daytime functioning, and quality of life in israeli adolescents. Chronobiol. Int. 28, 338–343. doi: 10.3109/07420528.2011.560698
PubMed Abstract | CrossRef Full Text | Google Scholar
Van der Maren, South., Moderie, C., Duclos, C., Paquet, J., Daneault, Five., and Dumont, G. (2018). Daily profiles of light exposure and evening employ of light-emitting devices in young adults complaining of a delayed slumber schedule. J. Biol. Rhythms 33, 192–202. doi: 10.1177/0748730418757007
PubMed Abstract | CrossRef Full Text | Google Scholar
Wilson, J., Reid, One thousand. J., Braun, R. I., Abbott, S. M., and Zee, P. C. (2018). Habitual light exposure relative to cyclic timing in delayed sleep-wake phase disorder. Sleep 41:zsy166. doi: 10.1093/sleep/zsy166/5078613
PubMed Abstract | CrossRef Full Text | Google Scholar
Wittmann, Thou., Dinich, J., Merrow, M., and Roenneberg, T. (2006). Social Jetlag: misalignment of biological and social time. Chronobiol. Int. 23, 497–509. doi: 10.1080/07420520500545979
PubMed Abstruse | CrossRef Full Text | Google Scholar
Woelders, T., Beersma, D. G. 1000., Gordijn, G. C. M., Hut, R. A., and Wams, E. J. (2017). Daily light exposure patterns reveal phase and flow of the human circadian clock. J. Biol. Rhythms 32, 274–286. doi: ten.1177/0748730417696787
PubMed Abstract | CrossRef Full Text | Google Scholar
Wright, K. P., McHill, A. West., Birks, B. R., Griffin, B. R., Rusterholz, T., and Chinoy, E. D. (2013). Entrainment of the human circadian clock to the natural light-dark wheel. Curr. Biol. 23, 1554–1558. doi: 10.1016/j.cub.2013.06.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Yetish, G., Kaplan, H., Gurven, M., Woods, B., Pontzer, H., Manger, P. R., et al. (2015). Natural sleep and its seasonal variations in three pre-industrial societies. Curr. Biol. 25, 2862–2868. doi: 10.1016/j.cub.2015.09.046
PubMed Abstract | CrossRef Total Text | Google Scholar
Zeitzer, J. M., Dijk, D.-J., Kronauer, R. E., Brown, E. N., and Czeisler, C. A. (2000). Sensitivity of the man circadian pacemaker to nocturnal lite: melatonin phase resetting and suppression. J. Physiol. 526, 695–702. doi: x.1111/j.1469-7793.2000.00695.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Source: https://www.frontiersin.org/articles/10.3389/fnins.2019.00882/full
0 Response to "Dijk D Archer S N 2009 Light Sleep and Circadian Rhythms Together Again Plos Biology"
Postar um comentário